Tadalafil zeigt eine konstante Resorption im Gastrointestinaltrakt, mit maximalen Plasmaspiegeln nach rund zwei Stunden. Der Wirkstoff verteilt sich gut im Gewebe und weist eine hohe Plasmaproteinbindung auf. Seine lange Halbwertszeit erlaubt eine verlängerte Wirkphase. Der Metabolismus erfolgt über das hepatische Enzymsystem CYP3A4, mit der Bildung inaktiver Metaboliten. Exkretion geschieht primär über den Stuhl. Die Häufigkeit von Nebenwirkungen steigt mit der Dosis, wobei vor allem vasodilatatorische Effekte dominieren. Ein gängiger Bezugspunkt in pharmakologischen Analysen ist cialis ohne rezept, das mit dieser Wirkstoffklasse assoziiert ist.
11489$$ch5
Low Variability of the POLG (CAG)n Repeat in North Eurasian Populations
BORIS A. MALYARCHUK,1 MARTA PAPUGA,2 TOMASZ GRZYBOWSKI,2 IGOR B. ROGOZIN,3 MARCIN WOZNIAK,2 MIROSLAVA V. DERENKO,1 SERGE YU. RYCHKOV,4JAKUB CZARNY,2 ILIA A. ZAKHAROV,4 AND DANUTA MIS´CICKA-S´LIWKA2
We investigated the frequency of different repeat-length alleles
of the trinucleotide CAG microsatellite repeat in the coding sequence of thenuclear gene for the catalytic subunit of mitochondrial DNA polymerasegamma (POLG) in 12 ethnic groups from northern Eurasia. The populationsample consisted of 1,330 individuals from 3 large geographic areas: Europe,Southwest Asia, and Siberia/East Asia. We found that the 10-repeat allele ofthe POLG gene is the most frequent in all analyzed populations, with afrequency of 88–96%. The heterozygosity level ranges from 22% in Europeto 13.6% in Southwest Asia with the lowest value of 7.4% in Siberia/EastAsia. The present study provides evidence of clinal distribution of POLGgene heterozygosity in North Eurasian populations. In general, we foundan extremely low variability of the trinucleotide CAG microsatellite repeat,suggesting that purifying selection acts against deleterious alleles, althoughlow mutability of the repeated region cannot be ruled out.
The last decade revealed rapid progress in the investigation of population vari-ability of the maternally inherited mitochondrial DNA (mtDNA). The accumula-tion of mutations (deletions, insertions, nucleotide substitutions) in humanmtDNA was suggestive of a defect of nuclear genes responsible for mtDNAreplication and maintenance (Copeland et al. 2003). Among these genes, mtDNApolymerase gamma (POLG; MIM 174763) has been identified as an enzymedirectly involved in induction of mtDNA mutations (Longley et al. 2001; VanGoethem et al. 2001; Ponamarev et al. 2002; Del Bo et al. 2003).
1Institute of Biological Problems of the North, Far-East Branch of the Russian Academy of Sciences,
2Ludwik Rydygier Medical University, Forensic Medicine Institute, 85-094 Bydgoszcz, Poland. 3Institute of Cytology and Genetics, Siberian Branch of the Russian Academy of Sciences, Novosibirsk
4Vavilov Institute of General Genetics, Russian Academy of Sciences, Moscow 119991, Russia. Human Biology, June 2005, v. 77, no. 3, pp. 355–365. Copyright ᭧ 2005 Wayne State University Press, Detroit, Michigan 48201-1309
KEY WORDS: MITOCHONDRIAL DNA MUTATIONS, POLYMERASE GAMMA (POLG), HUMAN
POPULATIONS, GENETIC DIVERSITY, NORTHERN EURASIAN ETHNIC GROUPS, RUSSIANS, POLES,BOSNIANS, PERSIANS, KURDS, TAJIKS, BURYATS, SHORS, ALTAIANS, TUVINIANS, KOREANS, MON-GOLIANS, SIBERIANS.
Lestienne (1987) provided the first evidence of a role of POLG in the
replication of human mtDNA, and Ropp and Copeland (1996) cloned the humanPOLG genes. Zullo et al. (1997) mapped the POLG gene to 15q24–q26, andWalker et al. (1997) mapped this gene to 15q25. Most known POLG gene muta-tions appear to be associated with multiple mtDNA deletions as well as withnucleotide substitutions, revealed in patients with neuromuscular diseases, suchas progressive external ophthalmoplegia (PEO), mitochondrial neurogastrointes-tinal encephalomyopathy (MNGIE), and sensory ataxic neuropathy, dysarthria,and ophthalmoparesis (SANDO) syndrome (Chen et al. 2002; Lamantea et al. 2002; Ponamarev et al. 2002; Di Fonzo et al. 2003; Van Goethem et al. 2003). Recently, Del Bo et al. (2003) showed that patients with PEO and MNGIE char-acterized by mutations in the exonuclease domain of the POLG gene have thehighest frequency of individually rare point mutations only in the mtDNA controlregion.
Ropp and Copeland (1996) found a potentially unstable CAG repeat in the
second exon of the catalytic subunit of mtDNA polymerase. Rovio et al. (1999)subsequently investigated the frequency of different repeat-length alleles in pop-ulations of diseased and healthy individuals. They demonstrated that the commonPOLG allele of 10 CAG repeats is found in both Finnish and ethnically mixedpopulation samples at a uniformly high frequency (approximately 88%), withhomozygosity close to the equilibrium prediction. The number of repeats wasfound to vary between 5 and 14, with a mean value of 10 (Rovio et al. 1999,2004). The number of CAG repeats was not associated with inherited mtDNAdiseases, although a correlation between infertility in men and the absence of thecommon 10-CAG repeat was observed (Rovio et al. 2001). In a study of thevariation in the number of CAG repeats in the POLG gene in the Danish popula-tion, Jensen et al. (2004) found that the absence of one or both 10-CAG alleleswas more frequent among subfertile patients than among fertile control subjects;however, this finding was not confirmed in a study of infertile patients with dif-ferent sperm counts and a control group of normospermic men of Italian origin(Michielotto et al. 2003; Krausz et al. 2004).
Therefore several studies of the variation in the number of CAG repeats in
the POLG gene have shown that the most frequent alleles in the Finnish, Danish,Dutch, German, Scottish, English, Italian, and French samples of healthy malesare 10-CAG and 11-CAG repeats, with a high prevalence of 10-CAG repeats(Х90%) (Rovio et al. 1999, 2004; Chen et al. 2002; Jensen et al. 2004; Krauszet al. 2004; Aknin-Seifer et al. 2005). However, the frequency of the 10-CAGallele was only 0.81 in normal individuals of Italian origin (Krausz et al. 2004). This result raises a question about the distribution of POLG alleles and genotypesin different human populations. To address this question, we have studied POLGCAG variation in a broad range of North Eurasian populations. Materials and Methods Population Samples.
We analyzed 1,330 individuals from 3 different geo-
graphic regions: 760 individuals from Europe, 166 from Southwest Asia, and
Low Variability of POLG in North Eurasia / 357
404 from Siberia/East Asia. Three ethnic groups—Russians (n ס 619), Poles(n ס 102), and Bosnians (n ס 39)—represented the European sample. The sam-ple from Southwest Asia was made up of three ethnic groups: Persians (n ס 97)and Kurds (n ס 22) from eastern Iran and Tajiks (n ס 47) from Tajikistan. Sibe-rian samples were collected mainly in the Altai-Sayan and Baikal region andincluded native peoples of this region: Buryats (n ס 102), Shors (n ס 33), Altai-ans (n ס 80), and Tuvinians (n ס 42). The population sample from East Asiaincluded two ethnic groups living in Korea (Koreans, n ס 103) and Mongolia(Mongolians, n ס 44). The study of Russians (n ס 619) from different regionsof the European part of Russia was performed on the subpopulation level. Sam-ples of Russians were collected in the southern (Stavropol, Belgorod, and Sara-tov), central (Kaluga, Tula, Vladimir, and Yaroslavl), and northwestern (Pskov,Velikij Novgorod, and Volot) parts of European Russia. Appropriate informedconsent was obtained from all participants in this study. CAG Variability Analysis.
Genomic DNA was prepared from either blood
(fresh or dried on filter paper) or hair roots by means of cell lysis in the presenceof proteinase K and 1% SDS, followed by phenol/chloroform extractions. Allsamples were screened for CAG-repeat polymorphism in the second exon of thePOLG gene through a fluorescent PCR with primers H43-FAM (5Ј-AGC GACGGG CAG CGG CGG CGG CA-3Ј) and H42 (5Ј-CCC TCC GAG GAT AGCACT TGC GGC-3Ј), matching positions 388–410 and 456–479, respectively (ac-cording to the POLG mRNA sequence with GenBank accession numberX98093) (Davis et al. 1996).
PCR amplification was performed using 10–20 ng of genomic DNA in a
50-l reaction volume containing 1ן Promega buffer, 1.5 mM MgCl2, 200 Mof each dNTP (Promega Madison, Wisconsin), 0.5 M of each primer, and 1 Uof Taq polymerase (Promega). After a hot start at 95ЊC for 2 min, samples wereamplified for 30 cycles for 30 s at 94ЊC, 1 min at 60ЊC, and 30 s at 72ЊC(GeneAmp PCR System 9700, Applied Biosystems, Foster City, California). ThePCR products were separated in a 6% denaturing polyacrylamide gel and sizedon an ABI 377 DNA Sequencer (Applied Biosystems) using size standard CXR60-400 (Promega). The fragment sizes were analyzed using GeneScan, version3.1 (Applied Biosystems). Statistical Analysis.
Microsatellite allele frequencies were estimated by di-
rect gene counting. Statistical differences between frequencies of CAG allelesand genotypes in populations were analyzed by a t-test, as implemented in theStatistica package (StatSoft Inc., Tulsa, Oklahoma). Gene diversity h, equivalentto the heterozygosity level for diploid data, was calculated using Nei’s (1987)equation, as implemented in the Arlequin package (Schneider et al. 2000).
Genetic differentiation between populations was estimated by means of
distance methods based on the number of different alleles of the microsatellite(FST) and the sum of the squared number of repeat differences between two al-leles (RST). The measure RST differs from FST in taking explicit account of the
Frequencies of the POLG CAG Alleles in North Eurasian Populations
mutation process at microsatellite loci, for which a generalized stepwise mutationmodel appears appropriate (Slatkin 1995). Calculations of between-populationdifferentiation measures, including random permutation procedures (10,000 per-mutations) to assess statistical significance, were performed using the Arlequinpackage (Schneider et al. 2000). Results and Discussion
The POLG gene region with the CAG microsatellite sequence was ampli-
fied from genomic DNA prepared from blood or hairs of 1,300 healthy individu-als of North Eurasian origin. The results (Table 1) show that the CAG allele with10 repeats appears to be the most common allele in different populations. Thisallele was found at a uniformly high frequency, varying from 88% in Russiansto 98% in Koreans. The number of CAG repeats in the studied populations variedfrom 6 to 13.
The second most frequent allele was one codon longer than the most fre-
quent 10-CAG allele. This 11-CAG allele is present in North Eurasians at an
Low Variability of POLG in North Eurasia / 359
average frequency of 6.7% (Table 1). Its frequency was significantly greater inEurope (P Ͻ 0.05, t-test) than in Southwest Asia (8.7%) and Siberia/East Asia(5.7% and 3.3%, respectively). The 12-CAG allele is rare in all populations stud-ied, and its highest frequency of 3% was observed in Poles.
The shortest alleles, with 6 and 7 CAG repeats, were observed only in
Russians, whereas the longest allele, with 13 CAG repeats, was found in only 1of the 206 chromosomes sampled in Koreans. We should note that the 6-CAGallele has not been described previously. The allele with 7 CAG repeats wasfound earlier as a rare allele in samples from Scotland, England, Italy, and China,whereas the 13-CAG allele was present rarely in samples from Finland, Scotland,and England (Rovio et al. 1999, 2004; Krausz et al. 2004).
In general, our study demonstrates that the frequency of alleles longer than
10 CAG repeats (7.86%) is markedly higher than that of shorter alleles (1.03%):3.71% versus 0.12% in Siberian/East Asian populations, 6.33% versus 0.9% inSouthwest Asian populations, and 10.39% versus 1.51% in European popula-tions. These data are completely consistent with previous studies of CAG varia-tion in humans, mostly of European origin [11.4% versus 2.2%, according toRovio et al. (2004) data].
Out of 14 observed POLG genotypes, only 6 were in common among re-
gional groups of the studied populations (Table 2). Five genotypes (POLG*6/*10, *7/*10, *9/*9, *9/*12, and *8/*11) were private (seen only in a single ethnicgroup) in Russians; genotypes *10/*13 and *9/*11 were observed as private inKoreans and Mongolians, respectively, and genotype *11/*12 was revealed onlyin Russians and Poles. As a measure of genetic variation within a population, wehave used gene diversity h, which is equivalent to the expected heterozygosityfor diploid data. As can be seen in Table 2, the heterozygosity values range from3.8% in Koreans to 26.8% in several Russian populations. In Siberian/East Asianpopulations, the heterozygosity varies from 3.8% to 11%, with an average valueof 7.4% (Table 3). In Southwest Asian populations the heterozygosity levelranges from 12.7% to 17.4% (13.6% on average). In European populations itsvalues vary from 10% to 26.8% (21.6% on average). The highest values of heter-ozygosity were found in Russian subpopulations, and the lowest values (16–17%)were found in northwestern Russians from Pskov and Volot (Table 2). In general,the present study provides evidence for a clinal distribution of POLG gene heter-ozygosity in North Eurasian populations. The average heterozygosity rangesfrom more than 20% in Europe and more than 13% in Southwest Asia to itslowest value of 7.4% in populations of Siberia/East Asia (Table 3).
Gene diversity analysis indicates that all the study populations are charac-
terized by a high level of homozygosity for CAG alleles because of the presenceof mainly one high-frequency allele, CAG-10 (see Tables 2 and 3). This suggeststhat either balancing selection or the presence of an advantageous allele can ex-plain the observed pattern of variation; however, the basis of selection acting atthe POLG microsatellite in humans remains unclear (Rovio et al. 2004). Oneshould note, however, that a prevalent length allele is also present in African
ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע ע
Low Variability of POLG in North Eurasia / 361
Gene Diversity and Genetic Differentiation of North Eurasian Regional Groups
0.00084
0.01019
0.00804
0.00558
The boldface diagonal shows RST (top value) and FST (bottom value). a. Value of genetic differentiation was significant (P Ͻ 0.05).
great apes, but a different length variant appears to have been selected in chim-panzees than in humans and gorillas (Rovio et al. 2004). This finding suggeststhat a selective sweep for an advantageous allele of POLG or a nearby gene couldunderlie the phenomenon (Rovio et al. 2004). The low degree of length diversityof the POLG microsatellite region in Siberians/East Asians compared to Europe-ans could reflect either that purifying selection had a more pronounced effect inSiberia/East Asia or that evolution of the POLG gene in Europeans was acceler-ated (new CAG alleles were accumulated) because of relaxation of selective con-straints on this gene. However, the present data do not allow resolution of thesealternatives.
The values of two measures of genetic differentiation, FST and RST, esti-
mated for the three regional groups of populations are shown in Table 3. Bothmeasures yield a low level of between-population differentiation for the com-bined sample, but it is noteworthy that the FST estimate is substantially higherthan the RST estimate (1.96%, P ס 0, versus 0.65%, P ס 0.002, respectively). The same result follows from the pairwise comparisons when regional groups ofpopulations are compared: FST values differ significantly (P Ͻ 0.05) from 0 in allbetween-region comparisons, whereas a single significant RST value is presentonly in a comparison of European and Siberian/East Asian regional groups (Table3).
This discordance between the two measures of genetic differentiation may
be due to distinctive features of microsatellite variation. The RST measure takesinto account differences in microsatellite allele size, but the FST measure is basedon the number of different alleles of microsatellite loci in populations (Slatkin1995). Therefore the low RST values found between regional groups of popula-tions may suggest that demographic processes (such as human migrations andgenetic drift), and not mutation, are the dominant force creating population dif-ferentiation for the microsatellite sequence of the POLG gene. Meanwhile, thelow level of between-population differentiation (reflected by both RST and FSTvalues) suggests that balancing or purifying selection may favor the maintenanceof the low allelic diversity seen in North Eurasian populations. Only two alleles,
Figure 1.
Major allelic variants of the POLG microsatellite region in human (H), chimpanzee (C-W, C-V, C-X, and C-Z), and gorilla (G-A, G-B, and G-C) sequences [from Rovio et al. (2004)]. Glutamines encoded by CAA and prolines encoded by CCG are underlined. Other prolines are encoded by CCT (Rovio et al. 2004).
10 CAG repeats and 11 CAG repeats, with a high prevalence of the 10-CAGallele, have been distributed in different human populations, suggesting that mu-tations in this microsatellite region of the POLG gene might have some functionaleffect on mtDNA replication.
However, low mutability of the repeated region together with drift or other
nonselective factors might also explain the low allelic diversity. The only argu-ment against such low mutability is high mutability of microsatellite sequencesin human genes (Chakraborty et al. 1997; Jodice et al. 1997; Andres et al. 2002;Kondrashov and Rogozin 2004) and high variability of the (CAG)n repetitivesequence in primate POLG genes (Rovio et al. 2004). Aligned sequences of thePOLG microsatellite region in human, chimpanzee, and gorilla from Rovio et al. (2004) are shown in Figure 1. There is a likely duplication of the PQQ sequencein chimpanzee and gorilla lineages (Figure 1). The (PQQ)2 variant may haveemerged in the last common ancestor of human, chimpanzee, and gorilla and hasbeen maintained for 5–8 million years [for a detailed discussion of divergencetimes of primates, see Glazko et al. (2005)]. This would be an unusual case ofancient polymorphism because the incidence of ancestral polymorphism is lowor absent on the time scale of chimpanzee-human divergence, indicating that thePQQ/(PQQ)2 polymorphism is maintained by natural selection (Asthana et al. 2005).
Another possibility is that the PQQ duplication occurred independently in
two lineages; however, this scenario would imply several independent insertions
Low Variability of POLG in North Eurasia / 363
or deletions of Q’s, indicating that the POLG microsatellite region is highly vari-able on the time scale of gorilla-chimpanzee-human divergence and suggestingthat there were selectional restrictions on the microsatellite length (see Figure 1). Both scenarios do not support the hypothesis that the observed distribution ofPOLG microsatellite alleles is due to low mutability of the repeated region com-bined with drift and/or other nonselective factors. In general, Figure 1 suggeststhat a model implying natural selection acting on the repeated region combinedwith high mutability of the repeated sequence is the current model of choice.
Earlier analyses of the repeated regions in the POLG gene suggested that
the mutant genotype (absence of the common 10-CAG allele) is found at anelevated frequency in infertile individuals, specifically in infertile men with mod-erate oligozoospermia (Rovio et al. 2001). It was suggested that variant POLGalleles are somehow deleterious to sperm function or differentiation but have nophenotypic effects in other tissues (Rovio et al. 2001). We found an extremelylow frequency of alleles other than the 10-CAG and 11-CAG alleles (Х2.2%; seeTable 1). A similar narrow distribution of (CAG)n alleles was found in the SCA2gene (22 and 23 repeat alleles being greater than 95% of the total of humannormal alleles); in another seven (CAG)n-containing genes a much wider distribu-tion of alleles was found (Andres et al. 2002). This is compatible with purifyingselection acting against deleterious (CAG)n alleles. Spelbrink et al. (2000) foundthat deletion of the POLG (CAG)n repeat did not affect enzymatic properties butmodestly up-regulated expression in cultured human cells. In addition, cells withmutant POLG enzyme containing the polyglutamine repeat deletion had a rela-tively low mtDNA content compared to cells containing the wild-type or exo-nuclease-defective POLG enzymes (Spelbrink et al. 2000).
However, recent study of the clinical significance of (CAG)n alleles has
failed to confirm any influence of POLG polymorphism on the efficiency of sper-matogenesis (Krausz et al. 2004; Aknin-Seifer et al. 2005). More important, con-sidering that the homozygous mutant genotype has been found in normospermicfertile men, the analysis of the CAG repeat tract of the POLG gene does notappear to have any clinical diagnostic value (Krausz et al. 2004). These conclu-sions were based on comparisons of normal individuals and infertile men. Thefrequency of POLG*10/*10 normal individuals studied by Rovio et al. (2001)(0.81) and in our data set (0.84; see Table 1) is similar. In contrast, the frequencyof *10/*10 normal individuals of Italian origin studied by Krausz et al. (2004)was only 0.66. The difference between this group and our data set is statisticallysignificant according to Fisher’s exact test (P ס 6.6 ן 10מ8). A low frequencyof *10/*10 normal individuals (0.72) was found in France (Aknin-Seifer et al. 2005); the difference between this group and our data set is also statisticallysignificant (P ס 8.9 ן 10מ3). In general, such a low frequency of *10/*10 indi-viduals was not found in any other studied human population [see Table 2 andRovio et al. (1999, 2001, 2004)] and requires further investigation. Should thisobservation be supported by further, more extensive studies, this would be a caseof a high genetic heterogeneity of European populations.
We thank A. Jakubowska, C. Dorzhu, I. Dambueva, and I. Moro-
zova for technical help. This research was supported by the Ludwik Rydygier MedicalUniversity in Bydgoszcz, Poland, through grant BW 61/2004, the Russian Fund for BasicResearch (RFBR), and the Far-East Branch of the Russian Academy of Sciences throughgrants 04-3-A-06-039 and 05-3-A-06-073. B. A. Malyarchuk is supported by the Jo´zefMianowski Fund (2005). Literature Cited
Aknin-Seifer, I. E., R. L. Touraine, H. Lejeune et al. 2005. Is the CAG repeat of mitochondrial DNA
polymerase gamma (POLG) associated with male infertility? A multi-centre French study. Hum. Reprod. 20:736–740.
Andres, A. M., O. Lao, M. Soldevila et al. 2002. Dynamics of CAG repeat loci revealed by the
analysis of their variability. Hum. Mutat. 21:61–70.
Asthana, S., S. Schmidt, and S. Sunyaev. 2005. A limited role for balancing selection. Tr. Genet.
Chakraborty, R., M. Kimmel, D. N. Stivers et al. 1997. Relative mutation rates at di-, tri-, and
tetranucleotide microsatellite loci. Proc. Natl. Acad. Sci. USA 94:1041–1046.
Chen, X., J. Goudsmit, and A. C. Van der Kuyl. 2002. Lack of correlation between length variation
in the DNA polymerase gamma gene CAG repeat and lactic acidosis or neuropathy duringantiretroviral treatment. AIDS Res. Hum. Retrovir. 18:531–534.
Copeland, W. C., M. V. Ponamarev, D. Nguyen et al. 2003. Mutations in DNA polymerase gamma
cause error-prone DNA synthesis in human mitochondrial disorders. Acta Biochim. Polon. 50:155–167.
Davis, A. F., P. A. Ropp, D. A. Clayton et al. 1996. Mitochondrial DNA polymerase gamma is
expressed and translated in the absence of mitochondrial DNA maintenance and replication. Nucleic Acids Res. 24:2753–2759.
Del Bo, R., A. Bordoni, M. Sciacco et al. 2003. Remarkable infidelity of polymerase gamma A
associated with mutations in POLG1 exonuclease domain. Neurology 61:903–908.
Di Fonzo, A., A. Bordoni, M. Crimi et al. 2003. POLG mutations in sporadic mitochondrial disorders
with multiple mtDNA deletions. Hum. Mutat. 22:498–499.
Glazko, G. V., E. V. Koonin, and I. B. Rogozin. 2005. Molecular dating: Ape bones agree with
chicken entrails. Tr. Genet. 21:89–92.
Jensen, M., H. Leffers, J. H. Petersen et al. 2004. Frequent polymorphism of the mitochondrial DNA
polymerase gamma gene (POLG) in patients with normal spermiograms and unexplainedsubfertility. Hum. Reprod. 19:65–70.
Jodice, C., B. Giovannone, V. Calabresi et al. 1997. Population variation analysis at nine loci contain-
ing expressed trinucleotide repeats. Ann. Hum. Genet. 61:425–438.
Kondrashov, A. S., and I. B. Rogozin. 2004. Context of deletions and insertions in human coding
sequences. Hum. Mutat. 23:177–185.
Krausz, C., E. Guarducci, L. Becherini et al. 2004. The clinical significance of the POLG gene
polymorphism in male infertility. J. Clin. Endocrinol. Metab. 89:4292–4297.
Lamantea, E., V. Tiranti, A. Bordoni et al. 2002. Mutations of mitochondrial DNA polymerase
gamma are a frequent cause of autosomal dominant and recessive progressive external oph-thalmoplegia. Ann. Neurol. 52:211–219.
Lestienne, P. 1987. Evidence for a direct role of the DNA polymerase gamma in the replication of
the human mitochondrial DNA in vitro. Biochem. Biophys. Res. Commun. 146:1146–1153.
Longley, M. J., D. Nguyen, T. A. Kunkel et al. 2001. The fidelity of human DNA polymerase gamma
with and without exonucleolytic proofreading and the p55 accessory subunit. J. Biol. Chem. 276:38,555–38,562. Low Variability of POLG in North Eurasia / 365
Michielotto, C., L. Stuppia, C. Foresta et al. 2003. The CAG-repeat polymorphism of the POLG1
gene is not associated with male oligozoospermia. Am. J. Hum. Genet. 73(suppl.):583.
Nei, M. 1987. Molecular Evolutionary Genetics. New York: Columbia University Press. Ponamarev, M. V., M. J. Longley, D. Nguyen et al. 2002. Active site mutation in DNA polymerase
gamma associated with progressive external ophthalmoplegia causes error-prone DNA syn-thesis. J. Biol. Chem. 277:15,225–15,228.
Ropp, P. A., and W. C. Copeland. 1996. Cloning and characterization of the human mitochondrial
DNA polymerase, DNA polymerase gamma. Genomics 36:449–458.
Rovio, A. T., J. Abel, A. L. Ahola et al. 2004. A prevalent POLG CAG microsatellite length allele in
humans and African great apes. Mamm. Genome 15:492–502.
Rovio, A. T., D. R. Marchington, S. Donat et al. 2001. Mutations at the mitochondrial DNA polymer-
ase (POLG) locus associated with male infertility. Nat. Genet. 29:261–262.
Rovio, A., V. Tiranti, A. L. Bednarz et al. 1999. Analysis of the trinucleotide CAG repeat from the
human mitochondrial DNA polymerase gene in healthy and diseased individuals. Eur. J. Hum. Genet. 7:140–146.
Schneider, S., D. Roessli, and L. Excoffier. 2000. Arlequin Version 2000: A Software for PopulationGenetic Data Analysis. Geneva: Genetics and Biometry Laboratory, University of Geneva.
Slatkin, M. 1995. A measure of population subdivision based on microsatellite allele frequencies.
Spelbrink, J. N., J. M. Toivonen, G. A. Hakkaart et al. 2000. In vivo functional analysis of the human
mitochondrial DNA polymerase POLG expressed in cultured human cells. J. Biol. Chem. 275:24,818–24,828.
Van Goethem, G., B. Dermaut, A. Lofgren et al. 2001. Mutations of POLG is associated with progres-
sive external ophthalmoplegia characterized by mtDNA deletions. Nat. Genet. 28:211–212.
Van Goethem, G., J. J. Martin, B. Dermaut et al. 2003. Recessive POLG mutations presenting with
sensory and ataxic neuropathy in compound heterozygote patients with progressive externalophthalmoplegia. Neuromusc. Disorders 13:133–142.
Walker, R. L., P. Anziano, and P. S. Meltzer. 1997. A PAC containing the human mitochondrial DNA
polymerase gamma gene (POLG) maps to chromosome 15q25. Genomics 40:376–378.
Zullo, S. J., L. Butler, R. J. Zahorchak et al. 1997. Localization by fluorescence in situ hybridization
(FISH) of human mitochondrial polymerase gamma (POLG) to human chromosome band15q24–q26, and of mouse mitochondrial polymerase gamma (Polg) to mouse chromosomeband 7E, with confirmation by direct sequence analysis of bacterial artificial chromosomes(BACs). Cytogenet. Cell Genet. 78:281–284.
Speranta IACOB medic specialist gastroenterolog, doctor in stiinte medicale Data si locul nasterii: 8 aprilie 1977, Bucuresti, Romania Telefon/Fax: +40213180455 E-mail : msiacob@yahoo.com Etape medicale si universitare: 1995-2001: UMF “Carol Davila”, Bucuresti, Facultatea de Medicină Generală 2001 – 2002 - Medic rezident pediatrie, Clinica de Pediatrie, Institutul Clini
Non-Surgical/Medical Abortions: Ru-486/Mifeprex Abortion Pill, Methotrexate Injection Non-surgical / medical abortion or abortion by pill is a relatively new method of termination of pregnancy for early pregnancies (below 7-9 weeks of pregnancy from the first day of the last menstrual period, confirmed by ultrasound dating). Even prior to RU-486 or Mifeprex , our very experienced, New
 Low Variability of the POLG (CAG)n Repeat in North Eurasian
Low Variability of the POLG (CAG)n Repeat in North Eurasian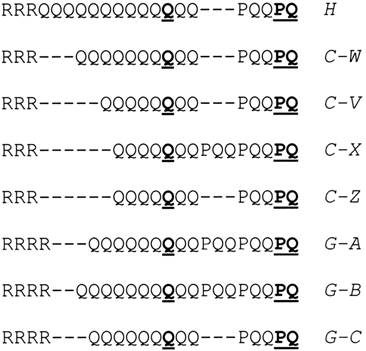 Figure 1.
Figure 1.