Tadalafil zeigt eine konstante Resorption im Gastrointestinaltrakt, mit maximalen Plasmaspiegeln nach rund zwei Stunden. Der Wirkstoff verteilt sich gut im Gewebe und weist eine hohe Plasmaproteinbindung auf. Seine lange Halbwertszeit erlaubt eine verlängerte Wirkphase. Der Metabolismus erfolgt über das hepatische Enzymsystem CYP3A4, mit der Bildung inaktiver Metaboliten. Exkretion geschieht primär über den Stuhl. Die Häufigkeit von Nebenwirkungen steigt mit der Dosis, wobei vor allem vasodilatatorische Effekte dominieren. Ein gängiger Bezugspunkt in pharmakologischen Analysen ist cialis ohne rezept, das mit dieser Wirkstoffklasse assoziiert ist.
Doi:10.1016/j.ibmb.2003.09.00
Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Identification of 1-lysophosphatidylethanolamine (C
antimicrobial compound in the housefly, Musca domestica
Karen Meylaers a,∗, Elke Clynen a, De´sire´ Daloze b, Arnold DeLoof a, Liliane Schoofs a
a Laboratory for Developmental Physiology, Genomics and Proteomics, Katholieke Universiteit Leuven, Naamsestraat 59, B-3000 Leuven 3010,
b Laboratory of Organic Chemistry, Free University of Brussels, CP 160/06, Av. F.D. Roosevelt 50, B-1050 Brussels, Belgium
Received 23 July 2003; received in revised form 4 September 2003; accepted 8 September 2003
Abstract
We observed that a methanolic whole body extract of uninfected last instar larvae of the housefly, Musca domestica, displayed
antifungal and antibacterial activity. We have further purified this extract to a single active fraction using reversed phase highperformance liquid chromatography. The pure fraction inhibited growth of the Gram-positive bacteria Bacillus thuringiensis andthe yeast Saccharomyces cerevisiae, but not the Gram-negative bacteria Escherichia coli. The active compound was determined tohave a molecular mass of 451.2 Da. Further analysis by nuclear magnetic resonance identified the substance as mono-unsaturated1-lysophosphatidylethanolamine (C16:1) (1-LPE). The structurally different and more common 2-LPE have been described asmediators of the antimicrobial activity of rimenophenazine antibiotic agents Our results suggest thatthe isolated 1-LPE displays a higher activity in comparison, possibly based on structure-specific differences in activity.
2003 Elsevier Ltd. All rights reserved. Keywords: Antimicrobial; Lysophosphatidylethanolamine; Musca domestica1. Introduction
induced only upon wounding and/or infection. As such,the reported purifications were mainly initiated from
The intensive application of antibiotic agents in health
immunised insects, meaning that the innate immune
care and animal breeding during the last decades has
response was induced by an experimental infection with
created a continuous selective pressure on micro-
pathogens or prepared fractions of these pathogens.
organisms. This has promoted the growth and spread of
Reports on constitutively expressed antimicrobial com-
microorganisms that display resistance against one or
several of the present antimicrobial agents. The develop-
ment of novel antimicrobial agents requires novel active
ever, that a constitutive expression and storage of anti-
lead-compounds. The innate immune system of insects
bacterial peptides occurs in granular cells of other invert-
has proven to be a source of a large variety of such com-
pounds and in particular of antimicrobial peptides
of non-peptidergic, low molecular mass compounds with
antimicrobial activity has, in comparison, been very lim-
Our preliminary experiments had indicated that a
whole body extract of uninfected larvae of the housefly,
In general, the expression of antimicrobial effectors is
Musca domestica, displayed growth inhibiting propertiesagainst bacterial cultures. Therefore, our goal was to iso-late
∗ Corresponding author. Tel.: +32-16-32-42-60; fax: +32-16-32-
E-mail address: karen.meylaers@bio.kuleuven.ac.be (K. Meylaers).
0965-1748/$ - see front matter 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.ibmb.2003.09.001
K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–492. Materials and methods
in 60 min; flow was 1 ml/min (3) analytical 4.6 × 250mm Suplex pkb-100 (Supelco); solvent A was 0.1%
aqueous TFA; solvent B was 0.1% TFA in CH CN; run
conditions: 0% B for 8 min then linear gradient to 90%
Pupae of M. domestica (Diptera, Muscidae) were
obtained from a local petshop. The flies that emergedwere fed on milk and dry diet (milk powder/sugar/yeast;2/2/1). Eggs could be collected from paper strips on
which they were deposited by the mature flies. Larvaewere reared on an artificial diet (milk powder/yeast/agar;
Growth inhibiting activity was determined in liquid
100/100/20 g per litre distilled water). When the larvae
growth medium using the Gram-positive bacteria Bacil-
are ready to start pupation, they will leave the moist diet
lus thuringiensis LMG 7138, the Gram-negative bacteria
and enter the sawdust that was spread on top, from which
Escherichia coli LMG 8063 and the yeast Saccharo-
they can then be collected. These larvae are referred to
myces cerevisiae ATCC 7754 as indicator organisms. B. thuringiensis and E. coli were grown in tryptic soy broth(Sigma) at 27 and 36 °C, respectively. S. cerevisiae was
2.2. Extraction and sample preparation
grown in Sabouraud maltose broth (Merck) at 27 °C. Overnight cultures of the bacteria and a 24 h culture
The last instar larvae were rinsed with distilled water,
of the yeast were diluted in fresh growth medium to a
dried on a paper towel and homogenised in a precooled
concentration of approximately 5 × 105 cells / ml. From
extraction solvent consisting of methanol/water/acetic
these dilutions, 10 µl inocula were taken and placed in
acid (90/9/1) complemented with 10 µg/ml of the pro-
the wells of a 96-well microtiter plate. Aliquots of the
tease inhibitor aprotinin (Sigma). After centrifuging the
HPLC samples to be tested were concentrated under vac-
homogenate at 13,000 × g for 30 min at 4 °C, the super-
uum to remove all traces of CH CN and TFA. The resi-
natant was collected. Methanol in the supernatant was
dues were dissolved in 40 µl suitable growth medium
removed by evaporation and the remaining residue was
and added to the inocula in the wells. Controls consisted
dissolved in 0.1% aqueous trifluoroacetic acid (TFA).
of 10 µl inoculum in 40 µl growth medium. Bacterial
Lipids were extracted by successively adding ethyl acet-
growth was evaluated after an incubation of 6 h, growth
ate and hexane, and each time removing the upper lipo-
rate of the yeast was evaluated after an overnight incu-
philic layer that formed after vigorously shaking. Traces
bation. The occurrence of growth inhibition was assessed
of these solvents were removed by short evaporation.
by visual observation using an inverted light microscope
The resulting sample was filtered through a 0.45 µm pore
(Nikon) and confirmed by absorbance measurement at
hydrophilic filter (Millipore) and loaded onto Mega-
were previously activated by flushing them with 50 mlof acetonitrile (CH CN), followed by 50 ml deionised
water. After loading the sample, the cartridges werewashed again with 50 ml deionised water. Next, theretained sample components were eluted sequentially
with 50 ml each of 30%, 60% and 90% CH CN in 0.1%
(ESI-Q-TOF) mass spectrometry was performed on a Q-
TOF system (Micromass). The purified active moleculewas identified by accurate mass analysis and fragmen-
2.3. High performance liquid chromatography (HPLC)
tation of the ion using collision induced dissociation(CID). Two microlitre of acetonitrile/water/formic acid
HPLC separations were performed on a Beckman
(80/19.9/0.1) containing the purified active compound
HPLC system with diode array detector. Chromatograms
was loaded in a gold coated glass capillary needle (L/Q
were constructed at 214 nm absorbance. The columns
needle, Proxeon) and sprayed from the needle with a
and run conditions applied for purification were success-
flow rate of 30 nl/min. Spectra were recorded in the posi-
ively: (1) preparative 25 × 100 mm Deltapak C (Waters
tive ion mode. Accurate mass measurements were per-
associates); solvent A was 0.1% aqueous TFA; solvent
formed by using the single protonated mass of riboflav-
B was 0.1% TFA in CH CN; run conditions: 0% B for
ine as a lock mass for internal calibration, which gives
6 min then linear gradient to 100% B in 90 min; flow
a mass accuracy of less than 10 ppm (parts per million).
was 40 ml/min (2) semi preparative 7.8 × 300 mm Xterra
Using the elemental mass software (Masslynx 3.5,
(Waters associates); solvent A was 0.1% aqueous
Micromass), any possible combination of prechosen
TFA; solvent B was 0.1% TFA in CH CN; run con-
atomic masses was calculated to match the measured
ditions: 0% B for 10 min then linear gradient to 90% B
mass within the predefined accuracy limits. K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–492.6. Nuclear magnetic resonance (NMR) analysis3.2. Identification of 1-lysophosphatidylethanolamine
A one-dimensional 1H NMR spectrum was recorded
A mass spectrometrical analysis indicated a mass of
in CH OD on a Varian Unity 600 apparatus (Varian Inc.)
at 600 MHz, using the residual O-H signal as internal
474.2), K+ adduct (m/z 490.2), K+H+ adduct (m/z 245.6)
calibration. The peak assignment was made by compari-
and dimeric ion (m/z 903.5) were also observed in the
mass spectrum, together with some fragment ions. Colli-sion-induced dissociation of the ion at m/z 452.2 resultedin several distinctive fragments, among which a domi-
3. Results
NMR analysis of the purified active molecule revealed
3.1. Antimicrobial activity in a larval extract of M.
a glycerol moiety, with the acyl chain positioned on the
second carbon of the glycerol backbone Although the position of the double carbon bond is not
An acidic extract of approximately 9300 last instar
undisputedly determined, it is most likely present in the
larvae (80 g) was prepared and loaded onto a solid phase
cartridge. Fractions, as eluted from the cartridge,
These data, together with the mass spectrometry data,
were tested for antimicrobial activity. The results are
identified the antimicrobial substance as 2-palmitoyl-1-
presented in Antimicrobial activity against B.
hydroxy-sn-glycero-3-phosphoethanolamine or 1-lyso-
thuringiensis and S. cerevisiae was found in the fractions
eluting with 60% and 90% CH CN in 0.1% aqueous
sponding molecular formula, C H NO P, was indicated
TFA. No antimicrobial activity was observed against E.
by elemental mass analysis within an accuracy of 5 ppm. coli up to a concentration of 30 larval equivalents.
The dominant ion at m/z 311.2 in the fragmentation
The antimicrobial activity in the fraction eluting with
spectrum is obtained after 1-LPE loses the phosphatidy-
60% CH CN in 0.1% aqueous TFA was lost upon further
lethanolamine group and corresponds thus to the acyl-
purification. The fraction eluting with 90% CH CN in
ated glycerol Further loss of the CH OH group
0.1% aqueous TFA was further purified on a preparative
from this fragment results in the ion at m/z 280.3.
reversed phase HPLC column (chromatogram not
Further loss of the glycerol from this fragment results in
shown). Two adjacent fractions inhibited growth of both
the ion at m/z 237.2 thus corresponding to the C
B. thuringiensis and S. cerevisiae. These fractions were
chain less OH. Also, a series of fragments, starting from
pooled and further purified on two different reversed
the ion at m/z 163.1 and ending with the ion at m/z
phase HPLC columns Antimicrobial activity
93.1, can be observed in the fragmentation spectrum and
was detected each time in a single fraction. After the
illustrates the repeated loss of a CH -group (14 Da) from
third purification step, a pure fraction was obtained. In
rough estimate, the quantity of sample necessary toinhibit growth of B. thuringiensis in our 50 µl liquidbioassay was calculated to be derived from an extract
4. Discussion
of 30 larval equivalents, all loss of material due to thesequential purification steps included.
We have isolated and identified 1-LPE in last instar
larvae of M. domestica, based on its growth inhibitingproperties against the Gram-positive bacteria B. thuringi-ensis and the yeast S. cerevisiae. The fraction containing
Antimicrobial activity of fractions as eluted from solid phase Mega-
1-LPE did not inhibit growth of the Gram-negative bac-
teria E. coli in the bioassay as described.
LPE are present in small quantities as integral
elements of membranes of animals, plants and bacteria. They are formed by hydrolysis of the structural mem-
brane component phosphatidylethanolamine (PE) byphospholipase A (formation of 1-LPE) and phospho-
lipase A (formation of 2-LPE). During the extraction of
uninfected larvae, the concentration of free LPE may
Saccharomyces cerevisiae ATCC 7754
have increased due to the degradation of cellular mem-
branes and the release of phospholipases. However, it
, Growth inhibition at a concentration of 1.25 larval equivalents; +,
2.5 larval equivalents; ±, 5 larval equivalents. Ϫ, No growth inhibition
should be considered that to a certain extent we are mim-
at a concentration of 10 larval equivalents.
icking the cell degradation that occurs in the course of
K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Purification of 1-LPE from a whole body extract of M. domestica on, subsequently, (1) semi preparative Xterra C
Suplex pkb-100. Run conditions as described in text. The arrows indicate fractions with antimicrobial activity. AUFS: absorbance units full scale. Absorbance at 214 nm is indicated by full line, absorbance at 280 nm indicated by dotted line.
an infection, and which will consequently also result in
mammalian cells for that matter, use diverse transport
an increase of LPE in the hemolymph, initiating a cas-
systems for K+-uptake and are thus not lethally affected
cade of hemolymph component interactions (further sub-
structural differences between mammalian and bacterial
The antimicrobial activity of rimenophenazine anti-
biotic agents was shown to be mediated by the release
of lysophospholipids. 2-LPE and lysophosphatidylcho-
become a basis for structure-guided development of new
line (LPC) have inhibited growth of Gram-positive bac-
teria with minimal growth inhibitory concentrations
Importantly, it should be noted that researchers gener-
) of 5–20 µg/ml, whereas Gram-negative bac-
ally refer to LPE or simply lysophospholipids in general,
teria are resistant at these concentrations
without specifying structural features. However, signifi-
This selectivity is apparently due to the
cant differences in activity of structurally different lyso-
selective inhibition of bacterial K+-transport systems, as
phospholipids are observed. Researchers have reported
that the observed haemolytic effect of LPC as well as
Gram-positive bacteria possess only a single
the phospholipase D (PLD) inhibiting effect of LPE dra-
operative K+-uptake system in normal growth con-
matically increase with the length and unsaturation of
ditions, whereas Gram-negative bacteria, as well as
the acyl chain and that only the intact LPE molecule is
K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Structural formula of 1-LPE and indication of some fragments
However, to exactly determine the hemolymph titre
and active concentration, we needed to obtain a quant-ified reference of 1-LPE. Unfortunately, the compoundas we purified it was not synthetically available andwould not be synthesized on request, as Avanti PolarLipids Inc. informed us, because the product is not verystable during chemical synthesis. The analogous 1-pal-mitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamine
Mass spectrometrical analysis of the antimicrobial HPLC
fraction on ESI-Q-oa-TOF mass spectrometer. (a) Mass spectrum (MS-
) was then ordered and tested in our liquid
mode). (b) Fragmentation spectrum (MSMS-mode) of the ion at m/z
growth inhibition bioassay. No activity against B. thurin-
452.2. Identified ions are indicated in italic superscript. M: original
giensis or S. cerevisiae was detected at concentrations
ion; M fr.: fragment of original ion. m/z: mass to charge ratio.
up to 125 µg/ml, in contrast to the report by However, growth inhibition in
the bioassay used by Van Rensburg was assessed
through the stop in bacterial uptake of radiolabeled
enhance superoxide responses in stimulated neutrophils
amino acids and is bound to be more sensitive than our
bioassay. Thus, this observation seems to confirm our
injury can be inhibited by antioxidants, but not the LPE-
suggestion that the growth inhibiting properties of our
isolated 1-LPE are stronger then those of the analogous
vations suggest that the specific structural configuration
2-LPE tested, as they were detected by our less sensi-
of the lysophospholipids is determinative for their effects
In addition to antibacterial activity, we observed a
Our data clearly indicated that the purified substance
growth inhibiting action of 1-LPE against the yeast S. cerevisiae. The antifungal action of 1-LPE remains
chain on the sn-2 position of the glycerol moiety. The
undetermined at the moment. Possibly, the general
reactive hydroxyl-group at the sn-1 position is thus much
reversible membrane-destabilising effect of the amphi-
more accessible in 1-LPE in comparison to its sn-2 pos-
pathic lysophospholipids may provide a basis for this
ition in the structural analogue 2-LPE, where it is par-
tially shielded by the acyl chain at the sn-1 position.
ture-specific action of 1-LPE must again not be
Therefore, it would be plausible that 1-LPE displays a
Finally, our data seem to be complemented by parallel
Table 21H NMR data of 1-LPE. Chemical shifts (d) are reported in ppm, coupling constants (J) in Hz, multiplicity (M) in description
CH–OCO
aliphatic CH
–CH –CH
–CH –CH –CO
HC–CH2–O–P
CH2–CH2–O–P K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
ongoing research in our laboratory on the immune
J., Bache`re, E., 1997. Penaeidins, a new family of antimicrobial
response of Drosophila melanogaster.
peptides isolated from the shrimp Penaeus vannamei (Decapoda). Journal of Biological Chemistry 272, 28398–28406.
demonstrated the upregulated presence of a PE-
Ginsburg, I., Ward, P.A., Varani, J., 1989. Lysophosphatides enhance
binding protein in larval hemolymph of D. melanogas-
superoxide responses of stimulated human neutrophils. Inflam-
ter, 25 min after induction of the immune response. In
Kakinuma, Y., 1993. K+ transport in Enterococcus hirae. In: Bakker,
that phospholipase A (PLA ) activity became upregul-
E.P. (Ed.), Alkali Cation Transport Systems in Prokaryotes. CRC
ated after E. coli infection of Sarchophaga peregrina.
Lamberty, M., Zachary, D., Lanot, R., Bordereau, C., Robert, A.,
The PLA -generated fatty acids, predominantly the eico-
Hoffmann, J.A., Bulet, P., 2001. Insect immunity. Constitutive
sanoid precursors, specifically participated in the acti-
expression of a cysteine-rich antifungal and a linear antibacterial
vation of the intracellular imd-pathway, leading to the
peptide in a termite insect. Journal of Biological Chemistry 276,
synthesis of antimicrobial peptides. One might speculate
that the hemolymph concentration of LPE becomes
Leem, J.Y., Nishimura, C., Kurata, S., Shimada, I., Kobayashi, A.,
Natori, S., 1996. Purification and characterization of N-beta-alanyl-
increased by the infection-induced upregulation of PLA-
5-S-glutathionyl-3,4-dihydroxyphenylalanine, a novel antibacterial
activity and/or by its release from damaged cell mem-
substance of Sarcophaga peregrina (flesh fly). Journal of Biologi-
branes and that these LPE subsequently bind to invading
microorganisms, thus causing not only the demonstrated
Leem, J.Y., Jeong, I.J., Park, K.T., Park, H.Y., 1999. Isolation of p-
microbial growth inhibition but also a further induction
hydroxycinnamaldehyde as an antibacterial substance from the saw
of the immune response by a signal transduction path-
fly, Acantholyda parki S. FEBS Letters 442, 53–56.
Mena, P.L., Djerassi, C., 1985. Synthesis of 5,9-hexacosadienoic acid
way that involves infection-induced (L)PE binding pro-
phospholipids.11. Phospholipid studies of marine organisms.
teins. Future analyses will further reveal the role of LPE
Chemistry and Physics of Lipids 37, 257–270.
Meylaers, K., Cerstiaens, A., Vierstraete, E., Baggerman, G., Michiels,
In conclusion, we propose that the specific structural
C.W., De Loof, A., Schoofs, L., 2003. Antimicrobial compounds
configuration of the isolated 1-LPE compound may
of low molecular mass are constitutively present in insects: charac-terisation of beta-alanyl-tyrosine. Current Pharmaceutical Design
enhance its antimicrobial reactivity in comparison to its
2-LPE analogue and provide a promising lead compound
Mitta, G., Vandenbulcke, F., Hubert, F., Roch, P., 1999. Mussel defen-
sins are synthesized and expressed in granulocytes, then releasedinto the plasma after bacterial challenge. Journal of Cell Science112, 4233–4242.
Otvos, L. Jr., 2000. Antibacterial peptides isolated from insects. Jour-
Acknowledgements
nal of Peptide Science 6, 497–511.
Relf, J.M., Chisholm, J.R., Kemp, G.D., Smith, V.J., 1999. Purification
This research was supported by the Flemish Science
and characterization of a cysteine-rich 11.5-kDa antibacterial pro-
Foundation (G0356.98 and G0187.00). Karen Meylaers
tein from the granular haemocytes of the shore crab, Carcinus
benefits from a scholarship from the Instituut voor de
maenas. European Journal of Biochemistry 264, 350–357.
Ryu, S.B., Karlsson, B.H., Ozgen, M., Palta, J.P., 1997. Inhibition of
aanmoediging van innovatie door Wetenschap en Techn-
phospholipase D by lysophosphatidylethanolamine, a lipid-derived
ologie in Vlaanderen (IWT-Vlaanderen). E. Clynen is
senescence retardant. Proceedings of the National Academy of
sponsored by the Fonds voor Wetenschappelijk Onder-
zoek in Vlaanderen (FWO-Vlaanderen).
Shigenaga, T., Muta, T., Toh, Y., Tokunaga, F., Iwanaga, S., 1990.
Antimicrobial tachyplesin peptide precursor. cDNA cloning andcellular localization in the horseshoe crab (Tachypleus tridentatus). Journal of Biological Chemistry 265, 21350–21354. References
Steel, H.C., Matlola, N.M., Anderson, R., 1999. Inhibition of potass-
ium transport and growth of mycobacteria exposed to clofazimine
Bulet, P., Hetru, C., Dimarcq, J.L., Hoffmann, D., 1999. Antimicrobial
and B669 is associated with a calcium-independent increase in
peptides in insects: structure and function. Developmental and
microbial phospholipase A2 activity. Journal of Antimicrobial
Comparative Immunology 23, 329–344.
Colles, S.M., Chisolm, G.M., 2000. Lysophosphatidylcholine-induced
Van Rensburg, C.E., Joone, G.K., O’Sullivan, J.F., Anderson, R.,
cellular injury in cultured fibroblasts involves oxidative events.
1992. Antimicrobial activities of clofazimine and B669 are
Journal of Lipid Research 41, 1188–1198.
mediated by lysophospholipids. Antimicrobial Agents and Chemo-
Daffre, S., Kylsten, P., Samakovlis, C., Hultmark, D., 1994. The lyso-
zyme locus in Drosophila melanogaster: an expanded gene family
Vierstraete, E., Cerstiaens, A., Baggerman, G., Van den Bergh, G., De
adapted for expression in the digestive tract. Molecular and General
Loof, A., Schoofs, L., 2003. Proteomics in Drosophila melanogas-
ter: first 2D database of larval hemolymph proteins. Biochemical
De Bruyn, E.E., Steel, H.C., Van Rensburg, E.J., Anderson, R., 1996.
and Biophysical Research Communications 304 (4), 831–838.
The riminophenazines, clofazimine and B669, inhibit potassium
Wrisch, A., Grissmer, S., 2000. Structural differences of bacterial and
transport in Gram-positive bacteria by a lysophospholipid-depen-
mammalian K+ channels. Journal of Biological Chemistry 275,
dent mechanism. Journal Antimicrobial Chemotherapy 38, 349–
Yang, J., Yamamoto, M., Ishibashi, J., Taniai, K., Yamakawa, M.,
Destoumieux, D., Bulet, P., Loew, D., Van Dorsselaer, A., Rodriguez,
1998. Isolation, cDNA cloning and gene expression of an antibac-
K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
terial protein from larvae of the coconut rhinoceros beetle, Oryctes
transgenic Drosophila reveals functional coupling between the
rhinoceros. European Journal of Biochemistry 255, 734–738.
phospholipase A2-generated fatty acid cascade and lipopolysaccha-
Yajima, M., Takada, M., Takahashi, N., Kikuchi, H., Natori, S., Osh-
ride-dependent activation of the imd pathway in insect immunity.
ima, Y., Kurata, S., 2003. A newly established in vitro culture using
The Biochemical Journal 371 (Pt1), 305–310.
ReminylTM ER Reminyl ER (extended release galantamine hydrobromide) As of June, 2006, Reminyl became available in the extended release (ER) format. It means that if you were taking Reminyl tablets twice a day prior to June, 2006, you would now take a Reminyl ER capsule once a day, in the morning. For example, if you are taking: • Reminyl 4 mg. twice a day, you would now take Reminyl ER
FOR A LIGHTER LIVING Das Grundprinzip von Rebalance wurde von Dr. med. D. Infanger, Leitender Arzt der Adipositassprechstunden des Stof wechselzentrums der Klinik Hirslanden entwickelt. es basiert auf mehr als zehnjähriger erfahrung mit über 2000 Patienten und wurde methodisch immer weiter verfeinert. Unser Kurs ist keine neue Wundermethode, sondern beruht auf dem aktuel sten Wissen aus schul

 Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Identification of 1-lysophosphatidylethanolamine (C
antimicrobial compound in the housefly, Musca domestica
Karen Meylaers a,∗, Elke Clynen a, De´sire´ Daloze b, Arnold DeLoof a, Liliane Schoofs a
a Laboratory for Developmental Physiology, Genomics and Proteomics, Katholieke Universiteit Leuven, Naamsestraat 59, B-3000 Leuven 3010,
b Laboratory of Organic Chemistry, Free University of Brussels, CP 160/06, Av. F.D. Roosevelt 50, B-1050 Brussels, Belgium
Received 23 July 2003; received in revised form 4 September 2003; accepted 8 September 2003
Abstract
Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Identification of 1-lysophosphatidylethanolamine (C
antimicrobial compound in the housefly, Musca domestica
Karen Meylaers a,∗, Elke Clynen a, De´sire´ Daloze b, Arnold DeLoof a, Liliane Schoofs a
a Laboratory for Developmental Physiology, Genomics and Proteomics, Katholieke Universiteit Leuven, Naamsestraat 59, B-3000 Leuven 3010,
b Laboratory of Organic Chemistry, Free University of Brussels, CP 160/06, Av. F.D. Roosevelt 50, B-1050 Brussels, Belgium
Received 23 July 2003; received in revised form 4 September 2003; accepted 8 September 2003
Abstract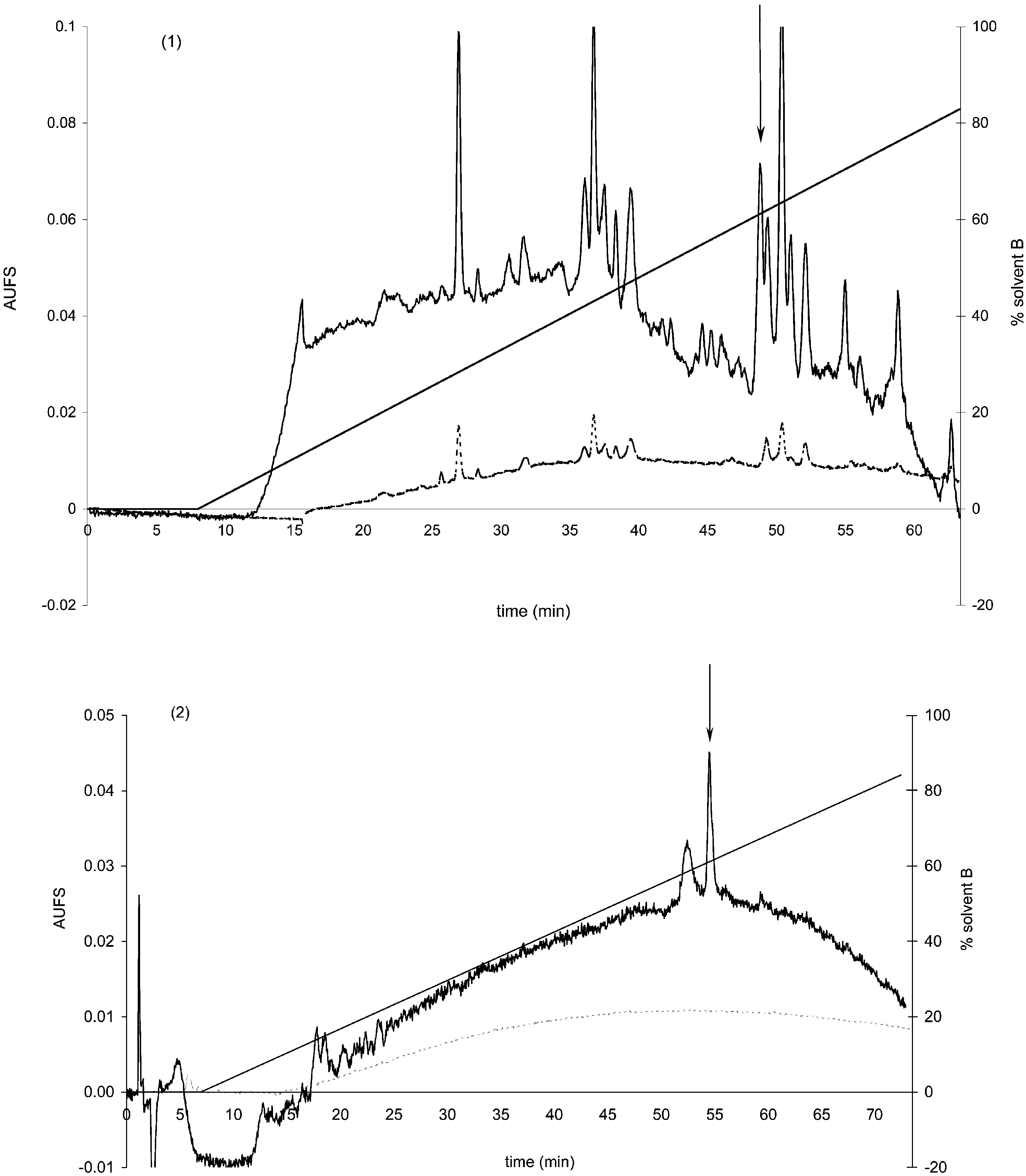 K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Purification of 1-LPE from a whole body extract of M. domestica on, subsequently, (1) semi preparative Xterra C
Suplex pkb-100. Run conditions as described in text. The arrows indicate fractions with antimicrobial activity. AUFS: absorbance units full scale.
K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Purification of 1-LPE from a whole body extract of M. domestica on, subsequently, (1) semi preparative Xterra C
Suplex pkb-100. Run conditions as described in text. The arrows indicate fractions with antimicrobial activity. AUFS: absorbance units full scale.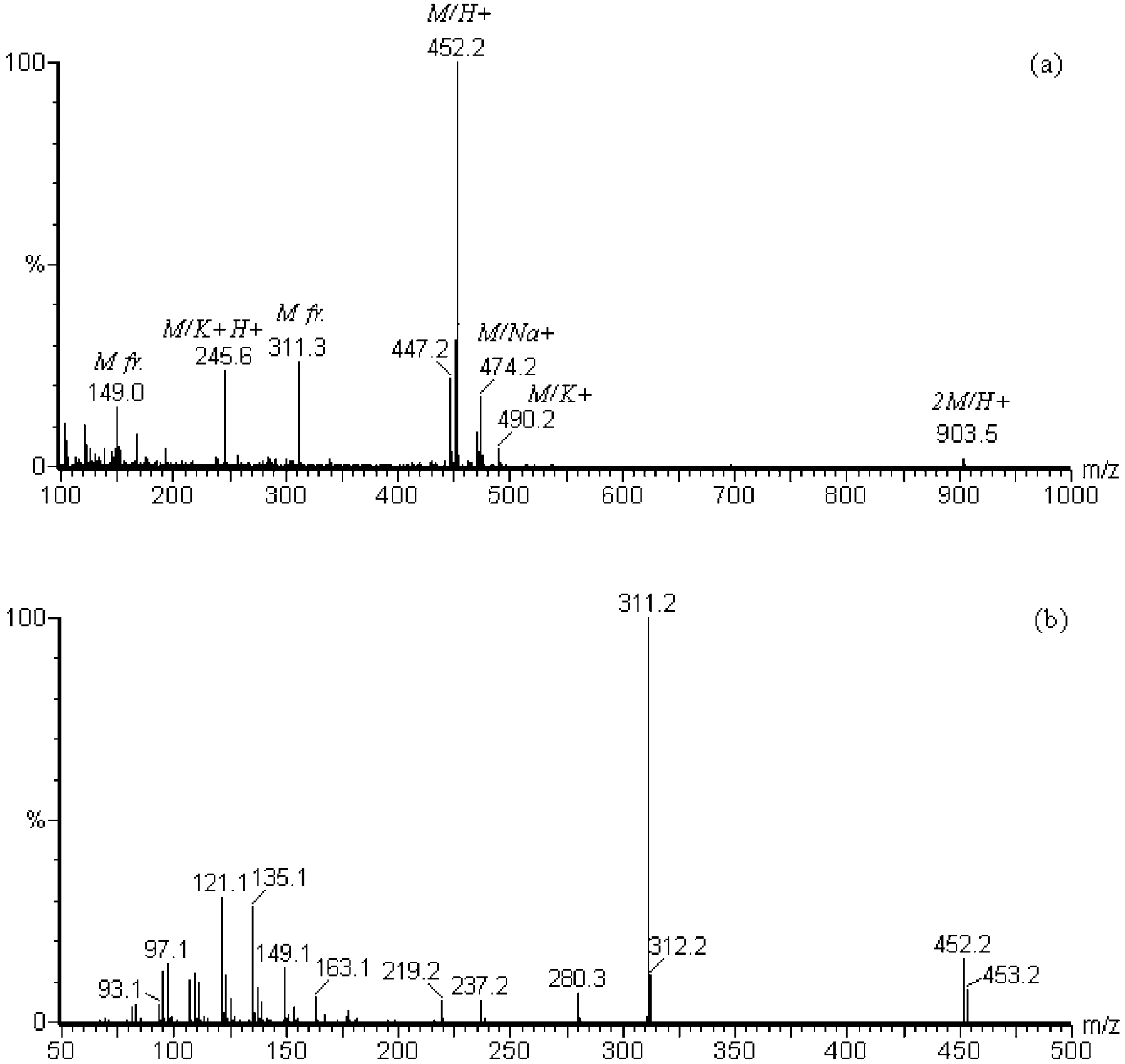
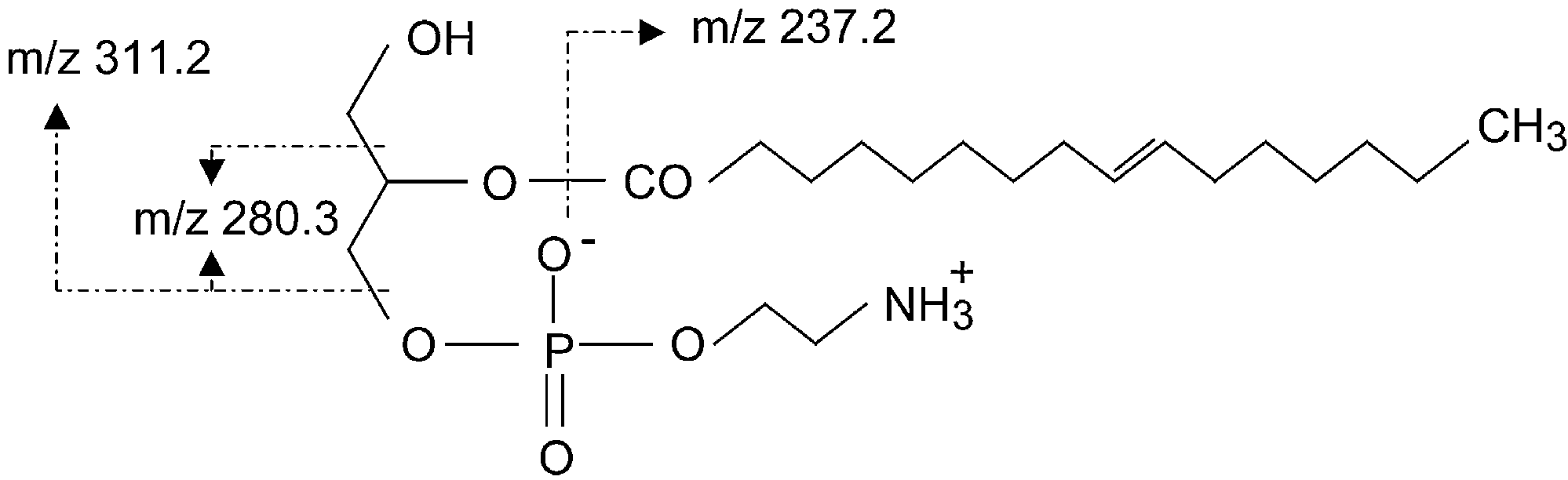 K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Structural formula of 1-LPE and indication of some fragments
However, to exactly determine the hemolymph titre
and active concentration, we needed to obtain a quant-ified reference of 1-LPE. Unfortunately, the compoundas we purified it was not synthetically available andwould not be synthesized on request, as Avanti PolarLipids Inc. informed us, because the product is not verystable during chemical synthesis. The analogous 1-pal-mitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamine
Mass spectrometrical analysis of the antimicrobial HPLC
fraction on ESI-Q-oa-TOF mass spectrometer. (a) Mass spectrum (MS-
) was then ordered and tested in our liquid
mode). (b) Fragmentation spectrum (MSMS-mode) of the ion at m/z
growth inhibition bioassay. No activity against B. thurin-
452.2. Identified ions are indicated in italic superscript. M: original
giensis or S. cerevisiae was detected at concentrations
ion; M fr.: fragment of original ion. m/z: mass to charge ratio.
K. Meylaers et al. / Insect Biochemistry and Molecular Biology 34 (2004) 43–49
Structural formula of 1-LPE and indication of some fragments
However, to exactly determine the hemolymph titre
and active concentration, we needed to obtain a quant-ified reference of 1-LPE. Unfortunately, the compoundas we purified it was not synthetically available andwould not be synthesized on request, as Avanti PolarLipids Inc. informed us, because the product is not verystable during chemical synthesis. The analogous 1-pal-mitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamine
Mass spectrometrical analysis of the antimicrobial HPLC
fraction on ESI-Q-oa-TOF mass spectrometer. (a) Mass spectrum (MS-
) was then ordered and tested in our liquid
mode). (b) Fragmentation spectrum (MSMS-mode) of the ion at m/z
growth inhibition bioassay. No activity against B. thurin-
452.2. Identified ions are indicated in italic superscript. M: original
giensis or S. cerevisiae was detected at concentrations
ion; M fr.: fragment of original ion. m/z: mass to charge ratio.